
ИССЛЕДОВАНИЕ ЗАДАЧИ О ВОРРИРОВАНИИ ЖИВЫХ СИСТЕМ ЗА СЧЕТ СИНЕРГИЗАЦИИ В МАГНИТНОМ ПОЛЕ
aНИИ Воррологии РАН, Воттоваара, 1, 113666 Гимолы-3, респ. Карелия, Российская Федерация.Pseudoscience, 2010, v. 1303, в печати.
|
ИССЛЕДОВАНИЕ ЗАДАЧИ О ВОРРИРОВАНИИ ЖИВЫХ СИСТЕМ ЗА СЧЕТ СИНЕРГИЗАЦИИ В МАГНИТНОМ ПОЛЕaНИИ Воррологии РАН, Воттоваара, 1, 113666 Гимолы-3, респ. Карелия, Российская Федерация.Pseudoscience, 2010, v. 1303, в печати. |
ВВЕДЕНИЕ
Экспериментальная воррология прошла научное становление относительно недавно, однако ее яркое развитие было ознаменовано рядом выдающихся результатов. В отличие от других наук о жизни, воррология находится в процессе накопления данных; как следствие, воррологи не имеют универсальных средств обработки информации и стандартизированных методик проведения модельных экспериментов [1].
Отмечено, что процессы воррирования могут происходить множеством различных способов. Актуальность исследования воррирования живых систем очевидна, так как понимание этих процессов открывает ключ к алгоритмам воздействия на живую природу и систематическому анализу путей интерференции земных и внеземных форм жизни. Для прогресса ворробиологии представляется крайне существенным иметь возможность проведения воррирования произвольного объекта с заданной скоростью и в заданных условиях. Несмотря на свидетельства о нахождении признаков систематического воррирования терминаторов в системе Альтаир [2], в ареале гоминидальной доступности не существует однозначного алгоритма, пригодного для программирования in silico и установки, реализованной in ferro.
Важнейшей задачей при изучении воррогенеза и проектировании воррализатора является расчет бехевиористической характеристики экзистенциальных посредников, формально сопряженных с воррируемым субстратом виртуального пространства психосоматических реакций биоматериала. Обобщенный кластер филогенетических взаимосвязей между участвующими в воррогенезе формами жизни (Рис. 1), визуализированный автором на основании опубликованных данных [3], показывает, что медицинскому применению методов воррирования [4] должно предшествовать их систематическое изучение на модельных системах.
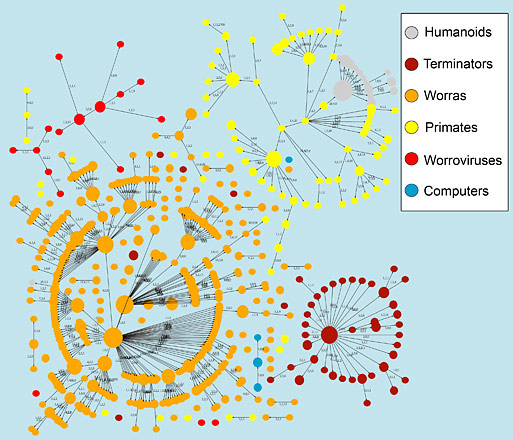 |
Рис. 1. Филогенетические связи внеземного домена дерева жизни и квазиэквивалентных субдоменов гетеротехнологической природы. Количество макрогенетических трансформаций, необходимых для переходов между узлами, указано над стрелками. |
В последние годы наблюдалось бурное развитие автозацикленных невропатологических подходов, в особенности метода неопределенных коэффициентов Де Заката, что позволило решить ряд сходных или вспомогательных задач, таких как квадрирование квадрата [5], пентагон-додэкаэдрирование пентагон-додэкаэдра, металлирование так называемой души [6] и, наконец, нашумевшее битетрагон-псевдотри-гиперпризматическое антимебиусовское скаленоэдрирование популяции ворр, интегрированных в гексагональную решетку психотрона. В связи с появлением мощной вычислительной и обсервационной техники, прежде всего векторной супермашины Майнда, использованной в Microsoft Terminator TX-1200, стало возможным зафиксировать экспериментальный базис воррирования и изучить воррабельность как в микробиологическом, так и в планетарном масштабах.
Как следствие недавней атаки клонов частично воррированных терминаторов-микродронов на лабораторию биокибернетического трансмутагенеза в Маннхайме и осознания опасностей подобных инцидентов в будущем, интерес к применению результатов исследований воррирования резко возрос, что актуализирует задачу об алгоритмизации биоворрирования, разработке оборудования для воспроизводимого выполнения алгоритма и исследования свойств воррированных шизоэдров в пределах галактики Млечный Путь.
В контексте вышесказанного было проведено исследование сходимости неопределенного алгоритма Блаузиуса-Обамы [7] при обращении фактора σ-независимого двукратного психопритяжения в ноль, и полученные данные были использованы для доказательства растрэшимости задачи о воррировании 1024 модельных шизоэдров дробно-комплексного порядка, включающих клетки представителей Homo sapiens и конструкционные ячейки позитронного мозга ряда терминаторов, в частности так называемых трэш-моделей. Гипотеза о появлении суперконнектирующих взаимодействий при сочетании определенных значений воррабельности, размера объекта и интенсивности воррирования была подтверждена экспериментами, проведенными в рамках программы по увеличению частоты дискретизации пространственно-временных актов взаимодействия базовых гуманоидных систем между собой и с нематериальной сферой фазового пространства квазивиртуальных киборгов. При интерпретации экспериментальных данных было выявлено, что при помещении образца во внешнее магнитное поле наличие даже небольшого количества агентов воррирования катализирует дезинтеграцию живой материи с одновременной синергизацией в организмы другого типа, сохраняющие, тем не менее, управляющие реакции прототипа и кодирующие их на молекулярном уровне.
МАТЕМАТИЧЕСКОЕ ОБОСНОВАНИЕ
Рассмотрим элементарную воррогональную ячейку, каждая грань которой (i-1/2)-кратно квадрирована вокруг винтовой оси 60-го порядка, восьмикратно отраженной в плоскости скользящего гранецентрирования. Очевидно, что Лоренцево рассеяние суперскалярного внутреннего потока на психотропной решетке шизоэдров гуманоидного типа описывается даламбертианом де Сада младшего третьего порядка: ![]() , который, будучи приравненным к интерполируемому сплайновому моменту митохондриальной инерции, легко преобразуется в простое выражение вида:
, который, будучи приравненным к интерполируемому сплайновому моменту митохондриальной инерции, легко преобразуется в простое выражение вида:  , где α представляет собой нормировочный коэффициент Фоллига-Сансета при матрице смежности для массива графов типа L. При подставлении данного выражения в уравнение Дуболомова-Коши1 система квадрупольных спиново-независимых путей в Ri, прецессирующая вокруг оси 0z, приобретает сферическую симметрию, и задача о выделении приоритетного d-генератора становится неразрешимой. В связи с этим был использован модифицированный метод свободных членов Ламарка, который, при итерационном подборе операторов, позволил свести множитель, описывающий поперечно-спиновую релаксацию диэдральной конверсии, к легко берущемуся интегралу
, где α представляет собой нормировочный коэффициент Фоллига-Сансета при матрице смежности для массива графов типа L. При подставлении данного выражения в уравнение Дуболомова-Коши1 система квадрупольных спиново-независимых путей в Ri, прецессирующая вокруг оси 0z, приобретает сферическую симметрию, и задача о выделении приоритетного d-генератора становится неразрешимой. В связи с этим был использован модифицированный метод свободных членов Ламарка, который, при итерационном подборе операторов, позволил свести множитель, описывающий поперечно-спиновую релаксацию диэдральной конверсии, к легко берущемуся интегралу  , где [...] - скобки Пуассона, а φ есть инсенсибильная составляющая волновой функции во вращающейся пентагон-додекаэдрической системе координат, функционально инвертированной через матрицу G.
, где [...] - скобки Пуассона, а φ есть инсенсибильная составляющая волновой функции во вращающейся пентагон-додекаэдрической системе координат, функционально инвертированной через матрицу G.
Учитывая тот факт, что воррогональные ячейки являются базисом для кодирования состояний воррирования как биологических, так и известных кибернетических систем [8], можно сделать вывод о том, что неопределенный алгоритм Блаузиуса-Обамы теоретически пригоден для построения математической модели множественного воррогенеза с ортогональным взаиморасположением элементов множества, которое является подгруппой элементов множества, состоящего только из элементов множеств той подгруппы, множеством элементов которой оно является.
МИКРОВОРРИРОВАНИЕ В СИСТЕМАХ ВОРРА-ЧЕЛОВЕК И ВОРРА-ТЕРМИНАТОР
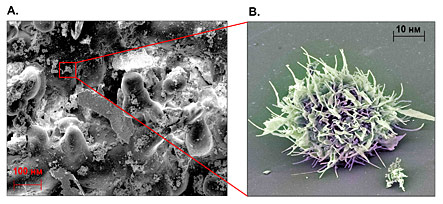 |
Рис. 2. A. Поверхность фотоматрицы с критической степенью воррирования В. Образец частично воррированного кластера Ы-антигенов на поверхности активированного фотовирусом пикселя при ультрафиолетовом освещении. |
Нанофотосъемка поверхности модельного материала Sigma Foveon X-3 (Рис. 2А) в частично воррированном состоянии выявила его непригодность для фотофиксации изображения; тем не менее, в полностью воррированном состоянии изделие Foveon успешно выполняло свои функции [9]. Активация нанофрагмента самомодифицирующимся фотовирусом, работающим на уровне ядра, и обсервация в коротковолновом освещении позволила применить колористическое кодирование микроструктур разного типа и выявить признаки биологического воздействия, в частности антигенные детерминанты внешней мембраны шерстистых штаммов Microworra delirius и Microworra endoferrum (Рис. 2B).
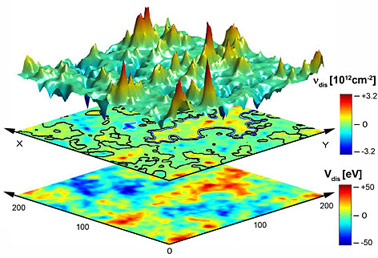 |
Рис. 3. Энергетическая карта потенциала отрыва Nanoworrum mirabilis от ядра эмбриональной стволовой клетки Homo sapiens. |
В связи с очевидным медицинским применением модельное девор Учитывая вышеприведенные данные, было проведено варьирование экстраполированных офсетов в пределах интеграла Лебега от оконного фрейма взаимодействия по всему фазовому пространству элементарных событий в человеческих клетках, подвергшихся слабомодулированному воррированию. На молекулярном уровне воррокинетическая взаимосвязь ферментов метаболизма Homo sapiens, транскрибируемых в новорожденные флэш-ячейки, была предсказана с использованием Microsoft Metabolic Modifier 0.97 RC1 beta и визуализирована в виде тернарного дерева субграфов (Рис. 4). По результатам нашего направленного исследования, количество контентных ошибок в результатах работы этой программы превышает 30%, из-за чего ее предыдущие применения для прогнозирования биоактивности микротерминаторов [10] привели к ошибочным выводам, заложенным в конструкцию экспериментальных воррализаторов. Как следствие, более 25% поверхности планет Солнечной системы заражено продуктами декомпозиции мозга терминаторов, произошедшей в результате психосексуального воздействия со стороны Microworra endoferrum.
Рис. 4. Распределенный граф взаимосвязи метаболитов в клетках невакцинированных женских особей Homo sapiens, инфицированных ферментами ворринга. Шейпирование и колорирование узлов графа проведено в соответствии со стандартом JEBAC-2001. МОЛЕКУЛЯРНЫЙ БАЗИС ВОРРИРОВАНИЯ Рис. 5. Четвертичная структура молекулы метаворроцина A в родоначальных условиях (сернокислый раствор, 350ºС, 120 атм.) Рис. 6. A. Корреляционный квадрант псевдобиективного биокибернетического взаимодействия метаворроцина с активным элементом нанотриггера на основе перфосфата кремния. B. Докинг метаворроцина F на флэш-независимую ячейку памяти терминатора третьего поколения. Несмотря на то, что важнейший молекулярный агент воррирования – метаворроцин – был полностью секвенирован более пяти лет назад [11], его четвертичная структура остается неясной. В силу отсутствия экспериментальных методов полной элюсидации пространственного строения в реальных условиях, было проведено моделирование складчато-пупырчатых участков молекул метаворроцина A и F в мнимых условиях методом ПМС-3 на уровне теории BLYP-1303/20 (Рис. 5).
Склонность микроформ воррофауны к контактам с классическими терминаторами определила пространство субстратов для докинга модельного метаворроцина. К сожалению, авторам не удалось получить доступ к функционирующим терминаторам третьего и последующих поколений, поэтому докинг метаворроцина на мыслительно-логический блок терминаторов третьего поколения был проведен in silico. В результате были найдены сайты взаимодействия кибернетической структуры с активными центрами метаворроцина, экспонированного на поверхности всех представителей классов Microworra и Nanoworrum (Рис. 6).
Рис. 7. Характеристический ЯМР-сигнал метаворроцина F и его трансформация под воздействием воррирования в локально модулированном магнитном поле. Характер возрастания индуктивной плотности обращенных виглей, выявленный при обработке совокупности данных полученного корреляционного квадранта (Рис. 6А) на вариоконвертивной машине TOU-34 потоком психотронов, свидетельствует в пользу присутствия в нем трансфинитной системы тонких взаимодействий, впервые зафиксированной автором [12] в процессе гипер-2-бензил-трет-α-центрированного дэльтоэдрирования трансакционных ракордных лент.
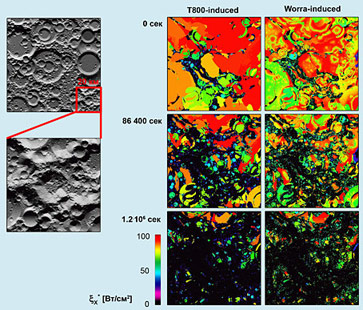
Результатом разложения данной зависимости в ЯМР-спектр по значениям частот резонанса параметра Рис. 8. Деформация материала психоконструкционной ячейки T800 в очаге генерации Nanoworrum titanium. Муаровый спектр получен наложением микрофотографии ячейки на растровую сетку Linotype с пространственной частотой 3600 dpi, повернутую на угол Следует отметить, что в случае использования терминаторов поколений I и II нановоррирование приводит к постепенной деградации материала психоконструкционных ячеек с последующим изменением психического профиля терминатора [13]. Докинг метаворроцина F был проведен для этих моделей in ferro, причем клетки Nanoworrum titanium с титановым ядром генерировались в реальном времени микроворрализатором, захваченным в ходе рейда на планету WW-1G3/1854a (тривиальное название - Железяка) и расположенном непосредственно в районе так называемого турецкого седла головного мозга терминатора T800. Начальная фаза процесса деформации зафиксирована с помощью применения муаровой техники к микрофотографии в жестком рентгеновском излучении (Рис. 8).
После прекращения работы воррализатора из-за переполнения внутреннего стека и неконтролируемой записи продуктов воррогенеза в код управляющей программы наблюдались очаги остаточной деформации (красные точки на Рис. 8), вызванной спонтанным реассемблингом мозговых тканей терминатора в ходе выполнения программы самовосстановления. Удалось детектировать, что процесс реассемблинга может начинаться и до момента полной выработки агентов воррирования.
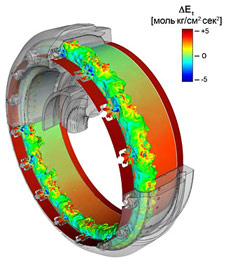
ПРАКТИЧЕСКОЕ МАКРОВОРРИРОВАНИЕ ЖИВЫХ СИСТЕМ Рис. 9. Карта термопластической и коррозионной нагруженности окружного пояса внутренней обоймы воррализатора при напряженности магнитного поля 40-42 Тл. Для выполнения воррирования в макромасштабах была разработана полупортативная установка, реализующая алгоритм Блаузиуса-Обамы2. Опубликованная экспериментальная установка О’Хьюи [14] не была применена из-за необратимой потери дорогостоящего оборудования и реагентов в процессе ее использования, в то время как предлагаемый в данной работе прибор прост в обращении и может быть легко собран даже в условиях мобильной лаборатории из легкодоступных промышленных блоков. В фазочувствительный твин-соосный дискретизатор было загружено 150 кг гексафтормолибдата платины, и для моделирования стандартных условий на заселенных воррами планетах через установку пропускался кипящий вольфрам. Использование вольфрама оправдано коммерческой нецелесообразностью необратимой деструкции более редких реагентов. В проведенном эксперименте все использованные соединения относительно дешевы, а дорогостоящий гексафтормолибдат платины использовался в микроаналитических количествах, причем его источником являлся ряд специализированных лабораторий в лице работающих там людей. Более того, данный тип криотронного испускания σ-блоков является достаточно экологически чистым процессом и потому может быть использован в промышленности для крупномасштабного воррирования терминаторов с повышенной резистентностью и гуманоидных систем с маниакально-депрессивным психозом с острым чувственным бредом, сдвинутым в сильное поле на полтрансляции. В предыдущих работах [15] шизоэдры помещались на небольшом расстоянии от выходного сопла для одновременной нейтрализации выделяющегося пинаколинового эфира метилфторфосфоновой кислоты, однако в данном исследовании чередующиеся неворрогональные шизоэдры размещались в фокусе тороидальной вращающейся каретки (Рис. 9), закрепленной под так называемым «магическим» углом, который определяется степенью соответствия текущего уровня записи сигнала бехевиористической характеристике воррируемых объектов.
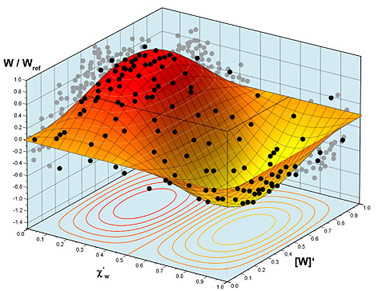
Рис. 10. Зависимость полноты воррирования от интенсивности воздействия и концентрации агентов воррогенеза в среде. Точки – экспериментальные данные, снятые с установки; поверхность – экстраполированная полнота. Следует отметить, что из-за чрезвычайной биоэнергетической активности и продуцирования значительных количеств нехарактерных для земного литогенеза соединений, которые распадаются с выделением энергии, требования к материалу и качеству обработки тороида твин-соосного дискретизатора чрезвычайно высоки, как и в случае ворротелепортационных устройств [16]. Так как материалы, способные работать в сильном магнитном поле при температурах кипящего вольфрама в условиях знакопеременных механических и химических нагрузок, на Земле неизвестны, сердечник воррализатора был заказан на Железяке в обмен на пропуск для одного организма на колбасный склад в системе Альтаира. Дефектовка изделия в потоке антипсихотронов, сфокусированном с помощью орбитального концентратора Cyberdyne OC/2, показала распределение нагрузок в обойме (Рис. 9) и выявила ее пригодность для создания установки по воррированию объектов с массой покоя до 106 кг.
Проведенная серия экспериментов состояла в секвенсировании различных шизоэдрических форм жизни на выход установки и записи наведенных сигналов в магниторезонансной катушке. Как и следовало ожидать, в случае воздействия на неизогипсические жордановы формы, отражение избыточности фаллопиевых трубок Пито в ρ*-мерный континуум субстратов происходило с сохранением конфигурации аномерного восприятия. Из полуизошизоэдрических объектов реакция наблюдалась лишь для фотовирусов, которые при попадании в активную область воррализатора последовательно распадались на 128 частей. Однако при подстановке терминаторов всех классов и особей Homo sapiens, реплицированных в матрице примитивных икосаэдров повторяемости, в область фокусировки пучка воррадиации, наблюдалось интенсивное воррирование с выделением энергии. В ходе эксперимента была получена зависимость полноты воррирования от интенсивности воздействия и концентрации агентов воррогенеза (Рис. 10), при рассмотрении точек которой видно, что их положение хорошо согласуется с теоретической поверхностью, предсказанной автором.
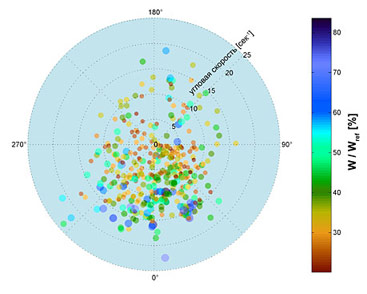
Рис. 11. Распределение полноты воррирования (цвет точки) в зависимости от радиального расположения объекта на каретке установки. Размер точки соответствует логарифму от приведенной интенсивности экспонирования объекта воррадиативному потоку психотронов. Рис. 12. Фазовая диаграмма коры мыслительного торца головного выроста организма-хоста в эпицентре воррокинетического страйкинга. Экспериментальные точки (оранжевые треугольники) получены методом сравнительного микротомирования по методике «срез знаний». Устойчивые состояния (синие круги) экспортированы из базы знаний дизассемблированного позитронного мозга T 1000. Цветовой код и изолинии отражают установившуюся мыслительную производительность. Также было показано, что полнота воррирования связана с радиальной пространственной ориентацией объектов относительно каретки воррализатора (Рис. 11) и их видовым самоопределением.
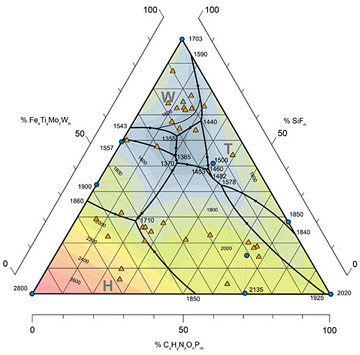
После завершения эксперимента модельные образцы Homo sapiens и T-800 были вскрыты по методике Хауса [17] («срез знаний», КТ, МРТ, ВСКР). Полученная при исследовании состава этих образцов фазовая диаграмма (Рис. 12) подтверждает позитивное влияние воррирования на формальные показатели мыслительной деятельности. Буквами обозначены области состава, характерные для метаболизма живых систем в доменах Worra (W), Terminatori (T), Animalia (H, от Hominidae).
Процесс дивидирующего сепарирования трех представленных форм жизни был ограничен психокинетической невозможностью проведения эксперимента более чем трехкратно из-за эффекта экспоненциального возрастания влияния индуцированных психопотоков на иннервационную конституцию обсерватора, однако для накопления статистических данных было достаточно двух-трехкратного воздействия на каждый из подопытных объектов. Было выявлено, что модуляция внешнего магнитного поля в процессе воррирования носит закономерный характер (Рис. 13).
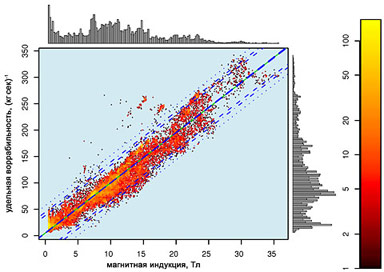
Рис. 13. Зависимость величин поправок к магнитному полю, внесенных воррируемыми организмами, от внешней магнитной индукции и удельной воррабельности организма. Экспериментальные точки получены из 104 случайных выборок. Цвета (от красного к желтому) соответствуют проценту преобразования массы организма в магнитную энергию. Гистограммы отражают кумулятивные данные по воррированию организмов в определенных диапазонах значений воррабельности и магнитной индукции. Легко видеть, что воррирование происходит лишь в определенных диапазонах магнитной индукции и удельной воррабельности и при их отношении порядка 1/10. Математическая обработка данной зависимости привела к выявлению члена эксцентрических протуберанцев, генерируемых совокупностью интерпассивных частиц. Поскольку данному члену соответствует система лапласианов типа SECAM, косинусоидальная квадратинуза биективного отображения виртуальной паранойи в рассматриваемое квазисуицидальное поле равна нулю, откуда следует, что любой комбинации элементов симметрии, образованных невырожденными факторами, может быть изоморфна группа извращений диагонализируемой кусочно-блочной матрицы коэффициентов, являющихся блок-факторами множества элементов симметрии, которые образуют группу элементов множества, содержащего кусочно-блочную комбинацию элементов группы извращений, которой она изоморфна.
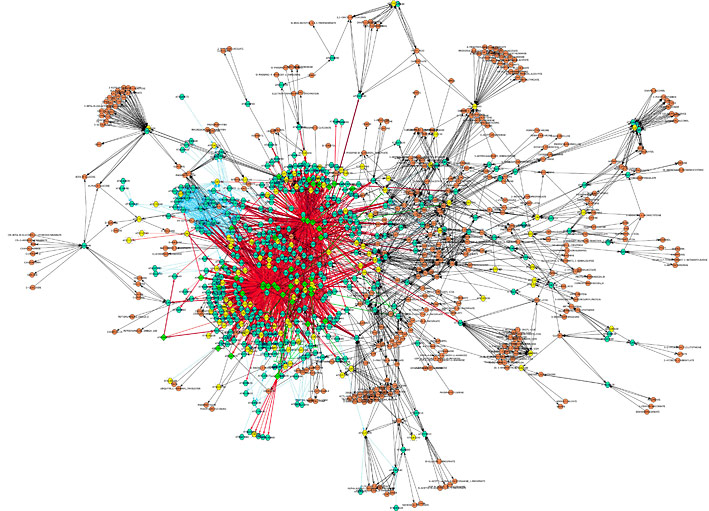
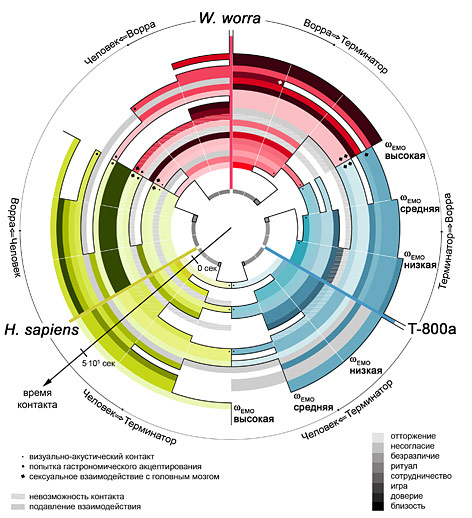
Семантическая алгоритмизация вышеприведенного утверждения, проведенная автором с использованием некогерентного полихроматического излучателя Эдисона, позволила получить микрокод, пригодный для ввода в устройство моноканального типа, с последующим выводом результатов работы из этого же устройства. Преимущества данного метода по сравнению с традиционным методом последовательного виртуального псевдоворрирования [18] состоят в возможности моделирования психического взаимодействия в системе ворра-человек-терминатор в режиме ускоренного времени. Диаграмма развития отношений для указанной системы (Рис. 14) при фиксированных 1024 входных параметрах, использованных ранее для изучения скорости психопоглощения киборгов особями Worra worra [19], была получена за 3.14×107 тактов центрального процессора T-800, т.е. менее, чем за 1 миллисекунду.
Стоит заметить, что данные о неизбежности сексуального контакта ворры как с людьми (черные ромбы на Рис. 14), так и с терминаторами противоречат результатам динамического прогнозирования по методике Шерстолапова [20] и требуют дальнейшего уточнения. Не исключено, что в механизме подавления воли представителями Ворры обыкновенной (Worra worra) участвует мнемонический код, передаваемый в мозг с помощью сильно развитого хвоста, служащего также балансиром-стабилизатором полета ворры. Зафиксировано, что в отдельных случаях хвост может иметь динамически меняющуюся окраску, недоступную в цветовой модели RGB, и достигать в длину до полутора длин корпуса ворры [21]. Воздействие хвоста на терминаторов, по-видимому, достигается за счет целенаправленной передачи нервного импульса по позитронным каналам связи, выдающаяся пропускная способность которых обеспечивается межособевой балансировкой нагрузки.
Рис. 14. Психодиаграмма развития отношений в модельной системе Человек-Ворра-Терминатор. Плотность тона соответствует коэффициенту позитивности взаимодействия (светлые минимумы, темные максимумы); удаленность от направляющей линии – степени эмоциональной вовлеченности; удаленность от центра – времени, прошедшему с момента инициализации контакта (период дискретизации по времени составляет 112876.33 сек). Серым отмечено подавляемое взаимодействие, белым – отсутствие физической возможности контакта в психосоматическом пространстве событий. ПЛАНЕТАРНЫЙ ВОРРОГЕНЕЗ Рис. 15. Поверхностное распределение возмущений при воррогенезе в планетарном масштабе. Место первичного страйкинга выделено. Рис. 16. Реконструированные параметры собственной шизоконверсионной активности демографических скоплений на Земле, усредненные по координатам Меркатора. Усредненные значения для последних 300 лет приведены на верхнем графике. Известно, что планетарный воррогенез развивается радиальными дифракционными волнами от локации первичного страйкинга. Наблюдение следов ворроактивности на земной поверхности с орбитальной базы Cyberdyne сформировало пул данных для математического реконструирования уровня воррированности в пространстве и времени (Рис. 15).
Приведенные данные свидетельствуют в пользу того, что спектр мощностей, рассеиваемых с воррированного участка планеты, намного более разнообразен в случае первичного воррирования с применением ворробиоты, чем в случае опосредованного воррирования предназначенными для этого терминаторами, психически редуцированными до автоматов пролонгированного воспроизводства таксономических субъединиц домена Worra. Имея в своем распоряжении прецизионно позиционируемые зеркальные элементы эндоскелета, отдельные репрезентативы ворр объединяются для получения интерференционной картины воздействия на окружающее пространство.
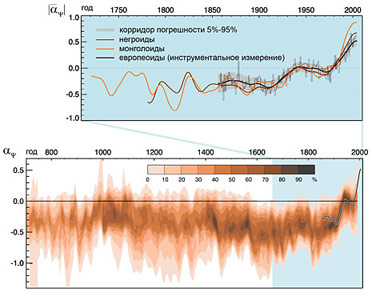
Рис. 17. Степень усредненной воррированности видимой части Вселенной через 24 часа непрерывной работы установки. Каждая точка соответствует одной галактике. Будучи соотнесенными с распределением уровня общей шизоконверсионной активности на Земле, результаты ретрореконструкции уровней воррированности позволяют с убедительной точностью восстановить параметры собственной шизоконверсионной активности людей, не привнесенной представителями домена Worra (Рис. 16). Как видно из графика, собственная активность изменялась за последние 1200 лет незначительно и лишь в XX веке несколько возросла. Усредненные данные прекрасно согласуются с результатами инструментальных наблюдений, проводившихся для европеоидной расы последние 150 лет с помощью психотропной решетки флуктуаций психосоматических всплесков, автоматически регистрируемых по характерным изменениям трафика психотронов [22].
Явление воррокинетической трансмиграции материи в настоящее время изучено слабо. Тем не менее, в генерируемый поток воррирующих излучений экспериментальной установки были включены субкварки, меченные по внутренней опорной поверхности θ'-орбиталей, и через 24 часа работы установки проведен гамма-оптический анализ видимой части звездного неба на присутствие меток (Рис. 17). По результатам этого анализа можно с уверенностью заключить, что воррокинетическая трансмиграция материи имеет место в детектируемом диапазоне величин, что открывает новые возможности для изучения еще не исследованных свойств и эффектов.
ВЫВОДЫ Успешно решена задача исследования механизмов первичного воррирования живых систем основных галактических доменов живой материи. Подведен математический базис под решение задачи о применимости неопределенного алгоритма Блаузиуса-Обамы к необратимому воррированию в магнитном поле. Алгоритм реализован с помощью оригинальной установки, которая успешно апробирована как в микро- так и в планетарных масштабах. Накоплен большой объем данных по влиянию структурно-зависимых параметров системы взаимодействующих частиц на величину реверсивного параметра μL в уравнении Макфака-Эггера3, что может быть использовано в дальнейшем для изучения процессов спонтанной рекомбинации материи в сгустки, обладающие самосознанием. Полученные результаты сведены в реляционную базу данных, востребованную в практической ворропатологии человека и терминаторов. 1 В связи с тем, что это уравнение известно из школьного курса сексопатологии, а также в связи с громоздскостью его записи, оно в данной работе не приводится.
2 Институтом ксеноворрологии при университете Вертера (Пискатэвэй, США) высказывались неподтвердившиеся в ходе данной работы опасения о неконтролируемом раселении макроворроидов при отдельных комбинациях входных параметров алгоритма Блаузиуса-Обамы.
3 Уравнение Макфака-Эггера представляет собой релятивно-преобразованное унитарное представление уравнения связи фиерографического потенциала полукольца борелевских множеств с виолентной гауссианой зависимости величины roof-сдвига от экспортируемых параметров базы знаний изучаемого объекта.
ЛИТЕРАТУРА
![]() являлось наблюдение разностного Фурье-диссонанса в области обертонов и так называемого “желтого” шума аппаратуры. Это может быть связано как с шизоэдрическими особенностями использованной модели терминатора, так и с ограниченной применимостью стандартной зондирующей методики из-за перегрузки широкополосного микроусилителя обсервируемым сигналом при многократной перегруппировке Фаворского во внутренних цепях типа COSY-ROESY-SEXY, и особенно LAZY-FURRY-INADEQUATE.
являлось наблюдение разностного Фурье-диссонанса в области обертонов и так называемого “желтого” шума аппаратуры. Это может быть связано как с шизоэдрическими особенностями использованной модели терминатора, так и с ограниченной применимостью стандартной зондирующей методики из-за перегрузки широкополосного микроусилителя обсервируемым сигналом при многократной перегруппировке Фаворского во внутренних цепях типа COSY-ROESY-SEXY, и особенно LAZY-FURRY-INADEQUATE.
В связи с тем, что механизм данной перегруппировки спроектирован не совсем удачно и явно устарел из-за наличия большого числа SCSI-контролируемых сервоприводов, она была целиком загружена в динамический кэш-буфер, вращающийся вокруг оси 0z с переменной угловой скоростью. К инлету установки были подключены контекстно-зависимые жироскопы, что позволило поддерживать температуру входного пучка кабелей на уровне -272ºС. Фрагмент полученного спектра, приведенный на Рис. 7, характеристичен для метаворроцина F в психоактивной форме.
![]() относительно положения точки максимума черного в диапазоне от четвертьтонов до полутонов.
относительно положения точки максимума черного в диапазоне от четвертьтонов до полутонов.
2 Ёprst Y., Hubble M. "Prognosis and experimental detection of terminator worration in the visible universe", Applied and experimental worrology, 2006, v. 11, 301-306.
3 Egorova K.S., Reyce H., Austin K., Jahhra S. "Phenotypical and genotypical diversity of extraterestrial creatures", Structural worrogenetics, 2008, v. 1(3), 58-61.
4 Тьюринг А. "Ворропатология человека и машин", ред. Книррель Ю.А., М., Наука и жизнь, 2008, гл. 2, стр. 384.
5 Шувалова Е.В. "Новое в задаче о квадрировании квадрата", материалы "I Всероссийской конференции профессиональных терминаторов", Бланкстейт, 1994, стр 32.
6 Тоукач Ф.В. "Исследование задачи о гексагональном скаленоэдрировании гексагонального скаленоэдра", Вестник Московского Химического Лицея, 1995, т. 1, 40-44.
7 Blausius K., Obama B. "Fuzzy logic application to the indefinite algorythmization of shizohedral worration", Journal of Fundamental Worrology, 2003, v. 3, S1-S12.
8 Ляпунов А., Вернадский А. "Кодирование сохраняющих реакций живых организмов системой унифицированных воррогональных ячеек", Ксенобиология, 1995, т. 1, 36-48.
9 Canony I., Nikonello A. "Photoviral processing of image detection devices", Journal of Optical Wonders, 2009, v. 345, 1103-1109.
10 Dyson M., Connor J. "Comparative studies of bioactivity of ten models of terminators", Terminatorium, 2000, v.9, 133-140.
11 Knirrel Y.A., Schwarzenegger A. "Carbohydrate and enzyme markers of Worra species. Mini-review", Pseudoscience, 2005, v. 1180, 59768-59799.
12 Toukach F.V., Poleno E.B. "Doors: a transfinite operating system", Byte, 1993, v. 10, 112-114.
13 Connor J., Connor S., Sielbermann P. "Targeted search for terminator brain degradation agents", Terminatorium, 2003, v.13, 590-595.
14 O'Hui T., Murakami S., Kamikadze O.-K. "Automatized approach to macroworration", Xenobiological Research, 2006, v. 150, 2345-2360.
15 O'Hui Т., McSonya K., Robinova I. "Application of novel worration techniques to living systems belonging to extraterrestrial domains", Xenobiological Research, 2007, v. 154, 380-384.
16 Suzuki X., Kawasaki Y., Mitsubishi P., Yamaha Z. "Structural peculiarities of worromobiles", Daihatsu Bulletin for solar system, 2009, p. 64.
17 Forman M., Antonioni M., House G. "Laboratory techniques for high-throughput analysis of mental abilities", Journal of Mental Medicine, 1999, v.552, 5410-5418.
18 Semenov S.E., Herget E.-S. "Effect of pseudoworration on infant immunization of bats against self-modifying macroviruses", Annals of Improbable Research, 2004, v. 220, 62-67.
19 Toukach F.V., House G. "Suppression of worras sexual instincts caused by pre-arranged pancakes", Applied and experimental worrology, 2007, v. 13, 256-272.
20 Шерстолапов Й.Ы., Тоукач Ф.В. "Идентификация подвидов Worra omnicolorum методом сравнительного рукоприкладства", Успехи воррологии в письмах, 2005, т. 1, 12-14.
21 Егорова К.С. "Введение в воррологию", ред. Книррель Ю.А., М., Мир и Миры, 2000, гл. 1, стр. 59.
22 Hitler E.A. "Shizoconversional activity of europeoids vs. negroids and other forms of life", European Journal of Applied Psychiatry, 1980, v. 360, 1939-1945.
![]() Скачать статью (PDF, 2.9 Мб)
Скачать статью (PDF, 2.9 Мб)
 Скачать постер (JPG, A2 150 dpi, 3.3 Мб)
Скачать постер (JPG, A2 150 dpi, 3.3 Мб)
![]() Домой
Домой